
For many people around the world, tonight is one of the most anticipated nights of the year. In that spirit, here’s some new science, not under the sun, but under the moonlight and starlight. New research from Wegener, Nagarajan and Pakrasi* describe something new about the Photosystem II D1 protein, an unexpected source that photosynthesis researchers had long written off as conquered or fully explained.
Why would we care about photosynthesis at night anyway? It all comes down to the distinctive lifestyle of the cyanobacterium Cyanothece, which belongs to an interesting class called unicellular diazotrophs (word of the day). Like all cyanobacteria, they perform oxygenic photosynthesis but they have the bonus biochemical ability of nitrogen-fixation.
Nitrogen-fixers are capable of pulling nitrogen gas (N2) out of the air and turning it into ammonia which the cells can use as a nitrogen source for important molecules like protein and DNA. Other microbes can perform nitrogen-fixation, and you may remember them as the root nodule symbionts of legumes. If you’re not a nitrogen-fixer, you have to rely on ammonia or other nitrate compounds present in the soil, sea or your diet (depending on what kind of organism you are) to make the required nitrogenous biomolecules. Since the Earth’s atmosphere is 78% N2, you don’t have to be great at math to figure out that acquiring the ability to tap into the atmospheric nitrogen source advances your species in the game of life a considerable number of squares.
What does this have to do with photosynthesis? Why don’t more plants and cyanobacteria get in on this nitrogen-fixation biochemistry? There’s a critical fatal flaw in the photosynthesis/nitrogen-fixation biochemical tango. Nitrogenase, the enzyme responsible for the nitrogen-fixation reaction is hopelessly and irreversibly killed by oxygen. Oxygen destroys the protein enzyme that’s already been made and tells the genome not to even bother wasting energy trying to make it new. If you’ve been paying attention to the blog, oxygenic photosynthesizers like cyanobacteria and plants make oxygen via their PSII enzyme every time light shines on them. So for nonphotosynthetic soil bacteria and other microbes, it’s not a big deal to avoid oxygen and go anaerobic to fix nitrogen. Cyanobacteria, however, must lead dangerous double lives to combine these two opposing biochemical processes.

Anabeana spherica via Wikimedia Commons The more round cell in the center of the string is the nitrogen-fixing heterocyst.
Some diazotrophic cyanobacteria solve this problem by separating these two processes in space. Species like Anabeana can go through a special type of development where the cells divide themselves, but incompletely to form a connected network like beads on a string. Every tenth cell (heterocyst) gets an extra layer of cell wall, destroys its photosynthetic machinery, and commits itself to a life of nitrogen-fixation. These cells share their fixed nitrogen compounds with their neighbors, which in turn share their fixed-carbon from photosynthesis. This is a fascinating process still hasn’t given up all of its secrets and remains an active area of research.

Cyanothece 51142 by Michelle Liberton, Pakrasi lab
Cyanothece, on the other hand, is a more ruggedly individualistic species that manages to perform both oxygenic photosynthesis and nitrogen-fixation within a single cell. It accomplishes this through a carefully controlled circadian rhythm. During the day, oxygenic photosynthesis occurs. During the night, nitrogen-fixation occurs. It sounds so simple, yet this elegance is far from easy. This is a true circadian process, in that, while some cues are definitely derived from light signals, the cells also have a strong internal clock that keeps the time of day and controls what biochemical machinery is active. Thus, near the hour of dawn and dusk, the cells are actively preparing for the metabolism to come. Also, if you were to train a culture of these cells under a certain day/night cycle for several days then switch them to continuous light, they would still maintain their circadian biochemical cycle for several days after the switch.
One issue that has puzzled researchers for some time is that the photosynthetic machinery, like PSII, is actively shut down at night. It’s not just that it’s dark, so there won’t be any photosynthetic oxygen production. Apparently, that’s not good enough. The cells actively shut it down. When researchers take culture samples of these cells during their dark period and shine light on them to check photosynthetic capability, there is no signal. Sure, there’s probably little chance any significant photosynthesis is occurring at night, but Cyanothece isn’t taking any chances. For these cells, no moonlight, floodlights from passing ships, comets, or even the blessed Eastern Star is going to interfere with their nocturnal nitrogen fixation.
Shutting down photosynthesis is no small feat. Again, if you’ve been paying attention to the blog, you will note that I’ve ranted on numerous occasions about how complex PSII is and how difficult it is to assemble and reassemble from its individual parts. As far as energetics are concerned, it would be a losing battle for Cyanothece to completely disassemble its PSII at dusk only to resynthesize it again the next morning. I don’t care how hard-up it is for a nitrogen source. As it turns out, Cyanothece and other unicellular diazotrophic cyanobacteria solve this problem in a more graceful way, for which Wegener et al have provided new experimental evidence.
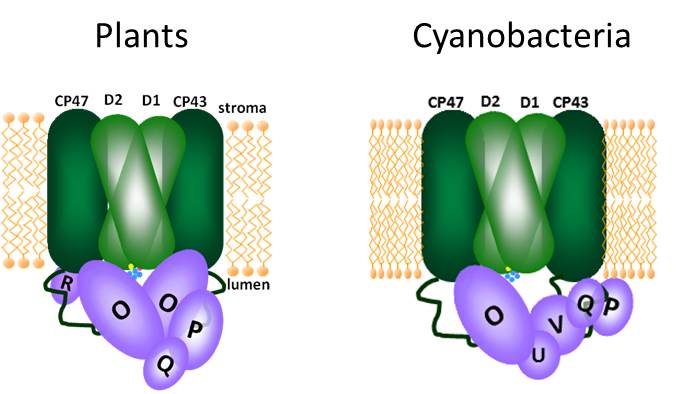
Cyanothece, like all other oxygenic photosynthesizers, contains multiple copies of the psbA gene encoding the D1 protein, which is a core subunit of PSII and critical for function. Since the advent of the genomic era, the genomes of numerous photosynthetic organisms have been sequenced revealing these multiple psbA genes, most of which have little or no protein sequence differences. These subtle changes have been linked to cells’ ability to fine-tune photosynthesis under certain conditions. However, Cyanothece and its cousins have a psbA gene copy that is particularly different- psbA4. If one were to take all of the site-directed mutants that photosynthesis researchers made to sort out PSII function over the last few decades since molecular biology techniques were available and threw them altogether into a single mutant gene, you would get psbA4. If PSII contained the D1 protein encoded by psbA4, there would be no C-terminal processing and no active site for water-splitting and oxygen production.
Why would any self-respecting photosynthetic organism retain such an utterly non-functional PSII subunit? The answer is because it does serve a critical function in the daily cycle of Cyanothece, just not during the day. The D1 protein encoded by the psbA4 gene, called a sentinel D1 protein (sD1), is made only for the night, when it does the only thing it still can do- fit right in the center of the PSII complex and prevent PSII function of any kind. There’s no chance of any oxygen production while sentinel D1 is on duty, but the cells can maintain most of the other PSII components in a stable complex (meaning they don’t have to be degraded and made fresh every morning). Then, Cyanothece can turn photosynthesis back on in the morning by replacing a single protein in its PSII complexes. The report by Wegener et al gives the first biochemical characterization of PSII complexes containing a sentinel D1 protein, providing evidence that this strategy works for controlling PSII oxygen production.

Performing oxygenic photosynthesis and nitrogen-fixation within the same cell is biochemically problematic, but the sentinel D1 protein appears to be the secret weapon for this dangerous double life. Through this strategy oxygenic photosynthesis and nitrogen fixation are “Always together, eternally apart.”** Many questions remain regarding the regulation of the transition from functional D1 to sentinel D1 at dusk and the reverse replacement at dawn. There are still outstanding questions regarding the D1 turnover event for any PSII in any photosynthetic organism, so it’s not at all clear what parts of this process Cyanothece uses or whether new factors are involved. Obviously, there must also be some sophisticated regulation to prevent the synthesis of sentinel D1 during the day.
While this work focuses on an organism you’ve never heard of, it has some interesting biotech implications for the future. As I mentioned, being able to both fix CO2 through photosynthesis and N2 through nitrogenase puts you at a significant metabolic advantage. And no, I don’t mean you. Remember, we’ve been through this- people are not photosynthetic. It would be great if one day we could engineer certain crop plants to combine these traits. Legumes already have this trick covered through symbiosis, but many other staple crops require tons of nitrogen fertilizer. If researchers could unlock the secrets of Cyanothece’s double life and translate it to plants, then it may be possible to engineer versions of crop plants that don’t require so much nitrogen fertilizer. Thus, one night in the future, engineered crop plants may be fixing nitrogen thanks to the addition of a variant gene they already had.
Johnna
*I know I’ve posted frequently about Pakrasi lab (my thesis lab) research, but this topic was just developing as I was leaving the lab and it’s too cool to pass up.
**Bonus points to you if you get the movie reference.
References and Links:














![Phycobiliproteins, bilin variation, and group III CA regulation. (A) Phycocyanin and phycoerythrin (blue and red lines, and in vials) absorb in regions of the visible spectrum not well absorbed by chlorophyll or carotenoids. Attached bilins: PEB, phycoerythrobilin; PCB, phycocyanobilin. (B) Natural diversity in coloration of many different cyanobacterial species due to variation in their bilin content [photograph by Christophe Six. Reproduced with permission from Six et al. (2007) (Copyright 2010, Biomed Central Ltd.)]. (C) Group III CA regulation model for F. diplosiphon in red light, showing the asymmetric regulation of red-light active genes (orange) and green-light active genes (yellow) by the Rca and Cgi systems. Dashed line represents proposed repression by the Cgi system; yellow balls, phosphoryl groups; blue boxes, RcaC binding sites.](https://newunderthesunblog.files.wordpress.com/2014/04/kehoepnas.jpg?w=561&h=528)














{kind=link}
{kind=link}