It has been a shamefully long time since I’ve done a post for the Journal Club category. So today’s will be a deluxe edition of Dickens-proportions. Normally, you only get the science tale as presented in any journal article, neatly fit to the scientific method. However, for every scientific publication, there is another tale, a more elaborate backstory with twists, turns and subplots. While these secondary tales may be more dramatic, the traditional publication process relegates them to the shelf locked inside lab notebooks. Well today you will be getting both tales, because I’ll be breaking down my latest accepted manuscript. Read the science version (Tale 1), the behind-the-science version (Tale 2) or both.
I’ll be enlisting the help of Charles Dickens because many of the quotes from A Tale of Two Cities are as true for the practice of science as they are for complicated human struggles with relationships, sacrifice and revolution.
Tale 1
“It was the best of times, it was the worst of times.” Charles Dickens
All science projects seem this way. They can begin so full of promise, then change direction down a pathway you were not expecting and perhaps did not want to follow. You may think you know what you are doing, but some results case doubts. You have come to a conclusion, but then do one experiment too many and it shatters.
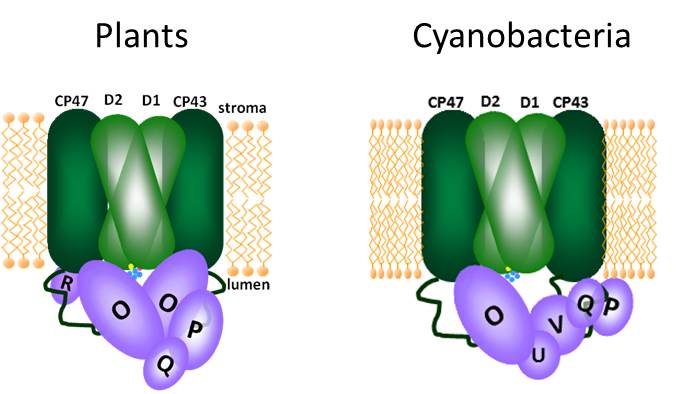
The PsbP-Domain Protein 1 (PPD1) Functions in the Assembly of Lumenal Domains in Photosystem I
Hypothesis: The lumenal protein PPD1 plays a critical role in photosynthesis, specifically in the accumulation of Photosystem I (PSI)
Experiments: In the model plant Arabidopsis, RNAi mutants of the PPD1 gene were characterized with respect to photosynthetic activity. The RNAi technique allows researchers to target a specific gene and knockdown its expression quickly and easily. These mutants can show a range of phenotypes that are useful in teasing apart the functions of genes whose complete elimination causes the death of the organism. The PSI activity in the PPD1 RNAi plants was extensively characterized as was the accumulation of many thylakoid membrane proteins (including PSI subunits). Native gel electrophoresis was also used to characterize the state of thylakoid membrane protein complexes in wild-type and PPD1 RNAi mutant plants.

PSI activity in PPD1 RNAi plants and representative plants from each group I-IV
Results: The PPD1 RNAi mutants with the lowest PPD1 expression were extremely small and pale green plants. Analysis of chlorophyll fluorescence showed that the mutants had much higher levels of fluorescence, indicative of an over-reduced plastoquinone pool and problems on the PSI-side of the photosynthetic electron transfer chain. Specific measurements of PSI activity showed that the PPD1 RNAi mutants had reduced amounts of active PSI reaction centers. However, energy could be transferred to these reaction centers by an alternative antenna system (LHCII). Moreover, the function of the PSI centers which did accumulate was not normal. Further analysis of protein accumulation in the thylakoids of the PPD1 RNAi mutants revealed there were specific problems in the accumulation of proteins on the lumenal side of PSI. In wild-type plants, the PPD1 protein was found to be associated with a thylakoid protein complex of ~300 kDa, which is smaller than any PSI complex.

2D gels showing thylakoid protein complexes in WT and PPD1 RNAi mutant. 1,2,4 complexes are forms of PSI; 3 is ATP synthase
Conclusions: The PPD1 functions in the proper assembly of PSI components on the lumenal side of the complex. In this area, PSI contains an extrinsically associated protein, PsaN, as well as extensive loop regions of the membrane proteins PsaA, PsaB and PsaF. All of these components create the binding environment for the soluble electron carrier, plastocyanin, which delivers electrons from upstream in the transfer chain. Reduced amounts of PPD1 affect the accumulation and assembly of these components. The mutant plants try to compensate for this loss of functional PSI by shifting some of the LHCII antenna such that it can funnel energy into PSI (the default for LHCII is to drive energy into PSII). The PPD1 protein is not considered a subunit because it was not found to be associated with fully assembled PSI complexes, but a smaller protein complex.
Think Ahead: The assembly of PSI is not a well-characterized biological process because, unlike PSII, PSI is an extremely stable enzyme. Thus, once it is assembled, the complexes can function properly for very long periods of time. Because it is a rare process, it is difficult to study. The original characterizations of PSI subunit mutants were performed many years ago, and it may be interesting to give them a fresh look with respect to PPD1 and some of the antenna effects we described. Identification of the other proteins in the complex with PPD1 (other PSI subunits perhaps) may help to define a PSI assembly intermediate. The secondary effect that the PPD1 mutation had on the antenna system will also be interesting to follow-up on because we don’t really know all of the details governing how plants allocate light energy between the photosystems. It is a sophisticated system with multiple layers of control.
Tale 2
“Nothing that we do, is done in vain. I believe, with all my soul, that we shall see triumph.” Charles Dickens
While scientific endeavors may have their dark moments, scientists tend to think that ultimately their research will see triumph. In the world of academia, this means publication in a peer-reviewed journal. Thus, all of the experiments that were done leading up to that publication but not included in it are not done in vain. They helped to work out the procedures necessary for acquiring the data that did appear in the figures. They were experiments that yielded negative data which eliminated hypotheses. Alas, those are never published.* It may be useful for scientists to know what wasn’t, but publishers only want to tell the stories of what was. (Hey, that almost sounds like Dickens too.)
The PPD1 story started with a blanket search for functions of the PPD family of proteins in the thylakoid lumen. They must be doing something to help plants photosynthesize, right? I was hopeful that maybe one of them had something to do with my favorite enzyme PSII. The way I chose to attack this problem was to characterize mutants of each of these proteins in Arabidopsis.
The easiest way to acquire Arabidopsis mutants is to order T-DNA lines (insertion mutants) for your gene of interest from the ABRC stock center. They send you seeds and you check to see if the mutants are useful or show a phenotype. There were two available lines (independent insertions) for the PPD1 gene and both of them were less than helpful to me. One line that another postdoc had been working with that had been passed on to me, which may have shown a phenotype, turned out to be heterozygous (not a complete mutant; still contains one wild-type PPD1 gene and should be normal). I couldn’t replicate any subtle phenotype, but neither could I find any homozygous (complete mutant) plants. Ever. I spent a lot of time verifying that that particular T-DNA mutant was embryonic lethal for homozygotes. Strange, but possibly extremely interesting. However, before taking this as a fact, it had to be confirmed that the homozygous lethal phenotype was because of the mutation in the PPD1 gene and not some other random mutation elsewhere in the genome. This can be sorted out by backcrossing heterozygote mutants to wild-type plants a few times and trying to recover the mutants. Ultimately, my experiments showed that the link between the PPD1 mutation and the embryonic lethal phenotype was not so absolute. At the same time, I was growing the other T-DNA mutant and it was proving to be equally unstable. Some plants would show a variegated phenotype, streaked leaves with patches of green and pale yellow. Other plants looked normal. I could never consistently link the phenotype to the ppd1 mutant genotype. With all of these inconsistencies in results, I decided that the T-DNA mutants were not useful in telling me anything about PPD1 function.
The alternative approach to Arabidopsis mutants is to use the RNAi technique to selectively suppress the expression of your favorite gene. While I was wrestling with the PPD1 T-DNA lines, I began the process of generating my own PPD1 RNAi lines. These plants turned out to be the most useful for figuring out what PPD1 does. When screening through these mutant individuals, there was a range in what the individual plants looked like- some looked almost normal while others were very small and pale. This is the good thing about RNAi because this range among individuals gives so a sort of picture of what is happening when the expression of the gene of interest is turned up or down over a gradient, like tuning up a brightness or volume knob on an old TV.
There was very little material to work with for the most severely affected PPD1 RNAi plants, but the biophysical measurements I could do on the tiny leaves indicated there was no problem with PSII. The defect was further downstream, probably in PSI. When using an instrument to specifically measure PSI function it was clear that was where the problem was. I would have to learn more about the PSI complex to say enough to turn my results into a publication, but at least I knew where this was going.
“There is prodigious strength in sorrow and despair.” Charles Dickens
The same week as my PSI results, I received an after-hours e-mail from my PI with the link to the following journal article: PsbP-domain protein1, a nuclear-encoded thylakoid lumenal protein, is essential for photosystem I assembly in Arabidopsis, Liu et al 2012 Plant Cell. When I quickly skimmed the abstract, my heart sank. My response was $%&^!, $%^@!, #$%@!, *&$%! I think I even drowned my sorrow in a pint of Hagen-Daz. There was only the slightest glimmer of satisfaction from the validation that researchers on the other side of the globe had come to the same conclusion as me.
Validation is not the name of the game. You see there is no prize for second place in scientific publishing. When you are the first group to publish a new idea, you have more control over the limits of the tale are. When you are second place, you cannot merely confirm what has been done (PPD1 has something to do with PSI). You must take it further, press on to unravel more details. Pressing on into the details of PSI territory was not really what I wanted to do.
However, after carefully reading what Liu et al had done, I reassessed my data and found a way to move forward. They had managed to characterize a clean T-DNA line, and the homozygous mutant plants they worked with were completely devoid of PSI. The small pale plants had to be grown on sucrose-containing medium since they could not support themselves photosynthetically. In my work, the RNAi lines allowed me to characterize plants that were very sick, but could still grow on soil. They accumulated some PSI, which could be analyzed more closely. Of course, that meant that I had to do a lot of experiments on precious little material. These experiments meant using a lot of brute strength just to get enough material for the experiments (spectroscopy measurements and blots, oh the blots!), investing time in fine-tuning protocols and money in antibodies for our second-favorite thylakoid complex.
“Vengeance and retribution require a long time; it is the rule.” Charles Dickens
Pushing forward with the experiments was difficult and took quite a bit of time. The sickest of the PPD1 RNAi lines were very small and would not set seed. Getting enough material meant screening for primary transformants every time. Learning the literature for a different enzyme complex was challenging. The papers describing the original characterizations of PSI subunit mutants were at least a decade old and often lacked data I would have liked to have seen. Not really flaws with that work; it’s just that what was not necessary for that work would have been extremely helpful to me.
Eventually, all of my data was written up in manuscript form and submitted away for peer review. I dabbled in other projects waiting the weeks until reviews came back. It took longer than usual, which meant only one thing- it was sent to a third reviewer. Yes, the two reviewers that initially evaluated my work had such differing views as to what my manuscript’s fate should be, a third reviewer was enlisted to help the editor in making the appropriate decision. Please revise with additional experimentation and there were specific concerns about how we went about doing some of our experiments.
Yes, a long time is the rule. I spent the next months painstakingly addressing the reviewer’s points with new experiments. One issue was how we estimated the amount of PPD1 protein in the mutants. With our antibody and a number of variations of our gel system, the PPD1 protein ran at the same molecular weight as the LHC proteins- the most abundant membrane protein on earth. These proteins obscured the signal for PPD1 such that we could never reliably estimate its amounts on denaturing gels. It either could not be seen or samples would require too much handling and treatment to consistently give a signal. However, PPD1 could be perfectly detected on native gels because the LHCs were nowhere near it in that system. Finally, I had point-by-point addressed all of the issues, revised the manuscript and created new figures.
We were ready to try again, but the tone of one of the reviewer’s comments gave us pause about resubmitting. Sure we had responses, but the original comments seemed like they would never invite satisfaction. There were some things about our results that would just not change. Experiments were done properly and yes, the results were still slightly unexpected. We would not be making up data because it would be easier for reviewers to accept. That is a cardinal sin in science, and a separate rule that should never be broken. For the revised manuscript, we took the chance on submitting to a separate journal with different reviewers for the chance of a favorable decision. This wager did not pay off because the two new reviewers had a completely separate set of comments to be addressed, many of which seemed impossible to satisfy with our sample limitations. We declined the invitation to revise and resubmit and ultimately resubmitted to the original journal. It went back to the original reviewers who seemed mostly satisfied with the improvements. Of course, there were some new comments by the reviewers that we could just not accommodate; not because the concerns were invalid, but because the questions went well beyond the scope of our work. There will always be more experiments to be done, but we firmly and politely stated we would not be addressing the new questions our latest experimental results sparked. We could only speculate as to future possibilities in the discussion section.
“A multitude of people and yet solitude.” Charles Dickens
In the case of this particular project, it was a multitude of data, yet not figures. I have numerous notebooks filled with raw data from experiments related to PPD1. In science, you go through a lot of preliminary work to get the answer, but when you show your work it must be much neater (certain colors, intensities, certain samples in certain orders). It’s like taking a math test and having a separate scratch paper (the lab notebook), but your answer is only a single circled number or neat graph (the manuscript figures). In this particular case, it was a multitude of blots. I cannot tell you how many blots I developed for this project. I practically lived in the dark room for months, eagerly waiting for films to emerge from the developer, praying the signals would be beautiful enough for figures.**
“I wish you to know that you have been the last dream of my soul.” Charles Dickens
Appeasing the reviewers in this final round felt a lot like the emotions in this quote. I had started forming the publication framework haphazardly because it wasn’t on a topic that I found exciting. Admittedly, I was only trying to do just enough to get acceptance. Even though my sentiment for some of my reviewers was more akin to a different saga, their requests did make the story better and forced me to expand my general knowledge on PSI and technical expertise in new protocols. All research can continue ever and on, but lines must be drawn somewhere because of the universal limits of effort, time and finances. I felt that the story was finally new and good with enough potential tangents to drive future research by possibly myself and others in the field. I had finally come to the point that I didn’t just want it done for the sake of adding another publication to the tally, but I wanted it published because the results deserved to be part of our body of photosynthesis knowledge.***
“It is a far, far better thing that I do, than I have ever done; it is a far, far better rest that I go to than I have ever known.” Charles Dickens
My PPD1 manuscript was eventually accepted at the Journal of Biological Chemistry this summer after a very long road of experimental struggles and research-related drama. Of all of my publications, this was definitely the most difficult to get to the point of publication. It is probably at the bottom of the list of my works if I had to rank them my favoritism based on any scale. This post was actually quite difficult to write; I so long to leave it in the past. However, I can now recognize that it is one of the better things I have done just to not give up on it. And after all the co-authors thought that everyone who would ever be interested in the PPD1 protein would have written the paper or reviewed it, we get an e-mail from another research group requesting our PPD1 antibody because their work may have a link to PPD1. “We read your paper with great interest,” they said in an e-mail sent a mere two days after our accepted manuscript appeared on-line. “They did!” I laughed. I sent them a sample, some behind-the-science instructions and well wishes. Apparently, my perseverance wasn’t just a better thing in terms of racking up publication numbers on my CV, but also for some other researchers embroiled in their own scientific epic. The best of times and the worst of times indeed.
As for me, it is a far, far, better rest that I go to than I have ever known as well. It’s not just a project change and definitely not the guillotine. Announcement coming soon to the blog.
Johnna
*However, Elsevier is launching a new journal where you can publish those results. Introducing the new journal New Negatives in Plant Science.
**There is a popular song this summer by Lil Jon and LMFAO “Shots”. There are not many lyrics. The rapper mostly just says shots over and over the backbeat track. To stave off insanity, or perhaps the opposite, I would sing my own version of the song “Blots.” All my biochemists, where y’at? Let’s go. When I walk in the lab, gloves on me, with the antibodies, I love chemiluminescence, I came to develop, lights off, it’s on! Blots, blots, blots, blots, blots, blots, blots, blots, blots, PPD1, blots, blots, blots, blots, blots, PsaB, blots, blots, blots, blots, blots, blots, LDS-PAGE, blots, blots, blots, blots, blots, Blue native, blots, blots, blots, blots, blots… If I don’t do these blots, I can’t resubmit! You get the idea. For visual effect, you can also picture me making it rain with x-ray films. Hey, what do you know another parody for the blog.
***Although if it would not have been accepted, I had threatened to just dump all of my results on this blog anyway and be done with it. Not sure how it ranks in terms of impact factor though.
References and Links:
http://www.jbc.org/content/early/2014/07/09/jbc.M114.589085
https://www.goodreads.com/work/quotes/2956372-a-tale-of-two-cities